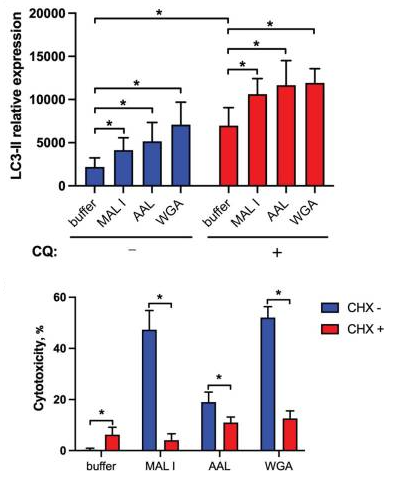
Cancer Immunology & Immune Modulation, Boehringer Ingelheim Pharma GmbH & Co. KG, Germanyらのグループは、骨髄性細胞に発現するC-Type lectin receptor 5A (CLEC5A) の発現と機能について報告しています。
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8896916/
骨髄性DAP12関連レクチン-1(MDL-1)としても知られるCLEC5Aは、病原体の表面に特異的に発現する糖鎖に優先的に結合する骨髄性Syk結合パターン認識受容体です。 CLEC5Aは主に骨髄細胞(単球、マクロファージ、好中球、樹状細胞)で発現し、IFN-γによってさらに発現が昂進されます。 CLEC5Aが結合するリガンドは、デング熱ウイルス、日本脳炎ウイルス、そしてまたA型インフルエンザウイルスに発現する糖鎖の末端フコースおよびマンノース構造として同定されました。更に、CLEC5Aは、細菌の細胞壁(リステリア・モノサイトゲネスや黄色ブドウ球菌など)の二糖類(N-アセチルグルコサミンおよびN-アセチルムラミン酸)に結合することも分かっています。機能的には、デングウイルスまたは他の病原体によって引き起こされるCLEC5A受容体の活性化は、炎症誘発性サイトカイン(TNF-α、IL-1、IL-6、IL-8、およびIL-17A)およびケモカイン(マクロファージ炎症性タンパク質-1α)、IFN-γ誘導タンパク質(IP-10/CXCL10)、およびマクロファージ由来ケモカイン(CCL22/MDC)の産生を誘導します。 CLEC5Aは、病原体関連抗原だけでなく、いくつかの内因性危険信号も認識し、その結果、無菌性炎症の病因に寄与する可能性があることも判明しています。
このレポートでは、単球由来マクロファージでのCLEAC5Aの発現状態と、α-CLEC5A AbによるCLEC5Aの選択的活性化による機能発現、および自己T細胞の活性化への影響が評価されています。
骨髄性細胞でのCLEC5Aの発現状態
CLEAC5Aの発現状態を幾つかの単球由来マクロファージ(MdM)間で比較しています。炎症性マクロファージ M1、中性マクロファージ M0、腫瘍形成性マクロファージ M2c、およびin vitro腫瘍関連マクロファージ(TAM)です。 CLEC5Aの発現は、単球および他のMdMサブセット(M0およびM2c)と比較して、M1 MdMで有意に上昇しましたが、TAMへの単球の分化はCLEC5Aの発現の低下をもたらしました。
CLEC5A活性化による機能発現
非感染性条件下でのCLEC5Aアゴニストの機能的効果を理解するために、M0 MdMのサイトカイン応答を評価しました。 α-CLEC5A Abに刺激されたM0 MdMにおいて、TNF-α、IL-6、IL-10、IL-1b、CCL22/MDC、CCL17/TARC、マトリックスメタロプロテアーゼ(MMP1)などのサイトカインおよびケモカインの産生が有意に昂進されました。興味深いことに、CLEC5Aの活性化は、CD80、PD-L1、CD206(MRC-1)、CD209(DC-SIGN)などの骨髄細胞特異的表面受容体の発現昂進をももたらしました。
最後に、無菌状態での骨髄細胞の選択的CLEC5Aを介した再プログラミングは、自家T細胞の活性化を促進するには不十分であるように見えました。