東北薬科大のグループは、病原菌認識に関わる11種の哺乳類レクチン受容体についてレビューしています。
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8185196/
このレビューの中から、3種のC-型レクチンについて、ご参考にまとめてみました。
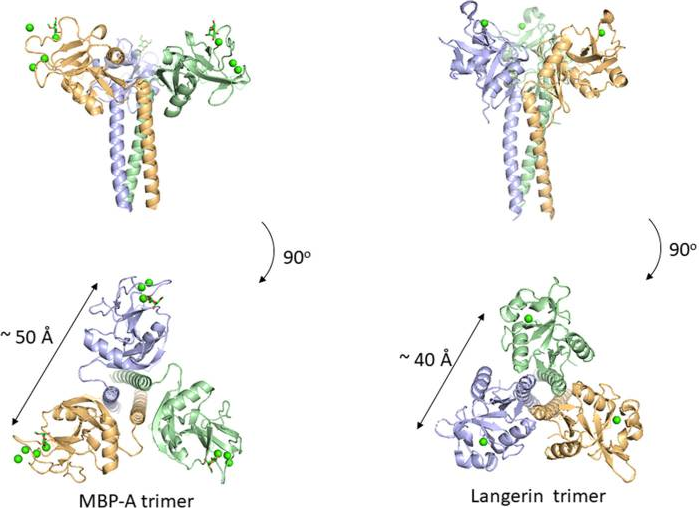
多くのC-型レクチンはマンノースに対して特異性を示し、病原体の認識に深く関わっています。一つ目のマンノース結合レクチンはMBP(MBLとも呼ばれる)ですが、これは自然免疫の補体経路を活性化するものとして知られています。マンノース修飾は哺乳類のN-型糖鎖で頻繁にみられるものであり、何故MBPが外因性のマンノースを認識できるのか不思議ではないでしょうか? 恐らくこれは、哺乳類のN-型糖鎖に比べて、病原菌のマンノース修飾は密度が高いことに原因があると考えられます。C-型レクチンのドメインは、マンノースのOH3とOH4に配位したCa2+イオンを好みます。そのアフィニティーはKd値が1mMと非常に低く、1:1の反応では免疫反応を引き起こせません。しかしながら、MBPは三量体であり、病原菌に発現する多くの末端マンノースと多価的に反応することができます。MBPの糖鎖結合サイト間の距離は50Åであり、そのような多価的な結合にちょうど良い構造となっています。もうひとつのC-型レクチンであるLangerinの場合には、三量体の結合サイト間の距離は40Åであります。 多価的に結合することによって、単一のマンノース結合に比べて、アフィニティーは何と1,000倍 向上することが分かっています。
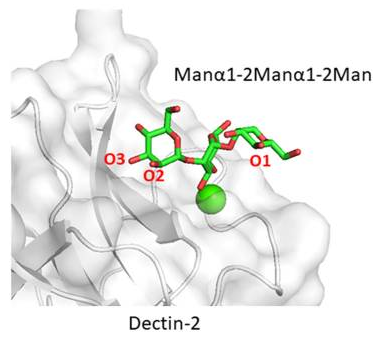
Dectin-2も同じようにC-型レクチンですが、DC-SIGN、Langerinなど他のC-型レクチンに比べると、Manα1-2Man 構造をまるっと認識することが異なっています。他のC-型レクチンは、末端のManα1-2Man構造しか認識しないのです。内部のマンノース残基を認識できるということは、Dectin-2 は長いマンナン型糖鎖を認識するに優れていると言えるでしょう。