UMR INSERM 1308 CAPTuR, Faculty of Medicine, University of Limoges, Limoges, Franceらのグループは、より正確にがん幹細胞を検出する新しい方法について報告しています。
https://www.nature.com/articles/s41416-024-02839-9
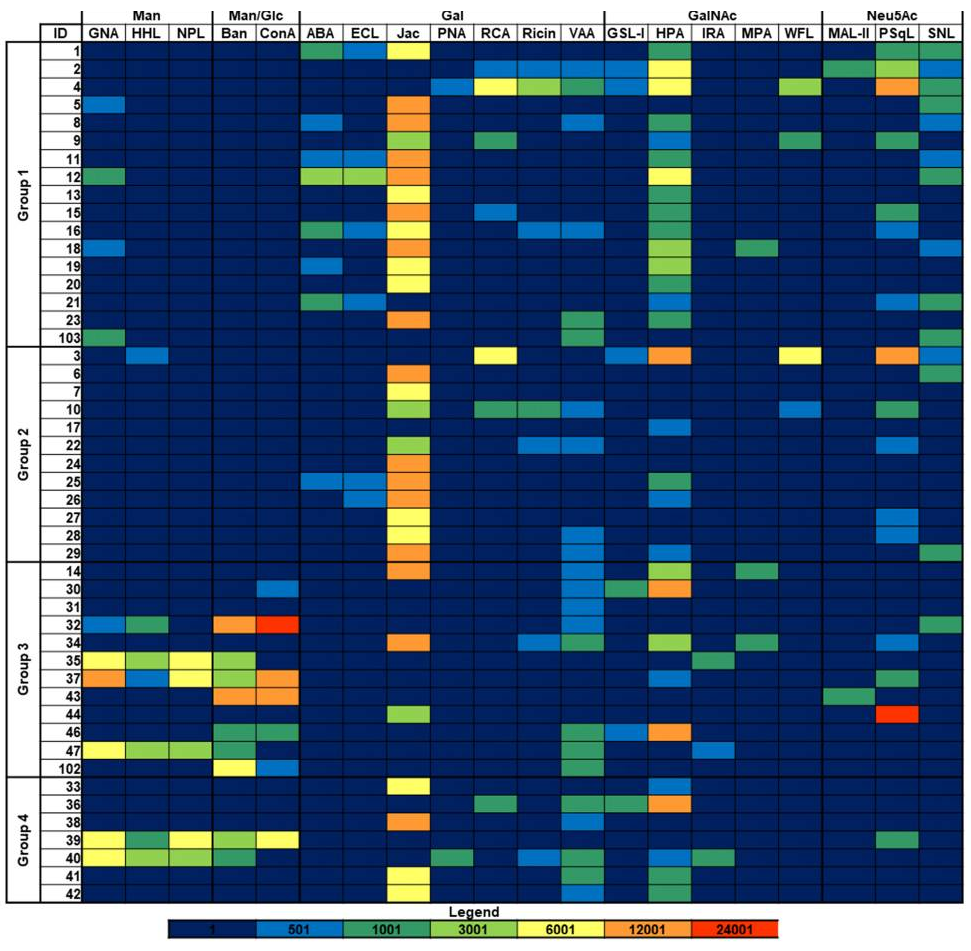
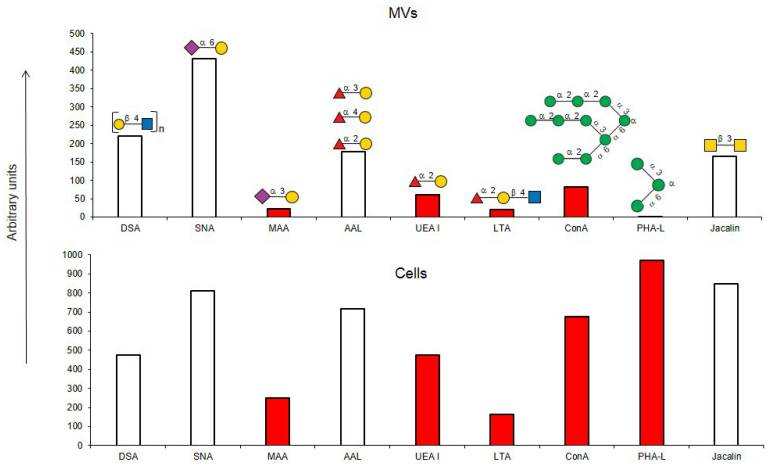
植物レクチンの組み合わせ (MIX: UEA-1 および GSL-1) が、不均一な非小細胞肺がん (NSCLC) 集団からがん幹細胞を検出するための新しいアプローチとして検証されています。
がん幹細胞上に発現した糖鎖修飾パターンを認識するレクチンの組み合わせは、CD133よりもがん幹細胞の検出と選別においてより効率的であることが実証されています。
CD133は既知のがん幹細胞マーカーとして知られています。