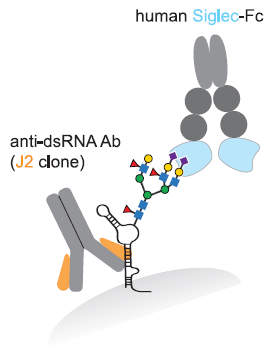
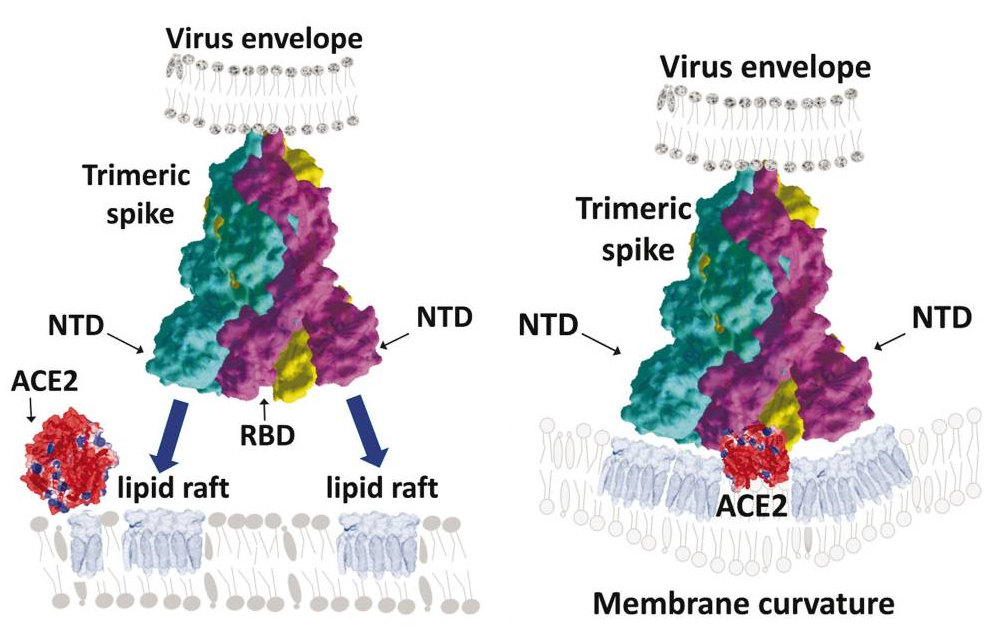
A group from Los Alamos National Laboratory, etc. has reported on effects of glycosylation onto binding between SARS-CoV-2 RBD and ACE2 with using molecular dynamics simulations.
https://www.mdpi.com/1999-4915/13/5/927
Two cases were evaluated: one is that six oligomannose (MAN9) glycans (magenta) are bound to ACE2 (at Asn53, Asn90, Asn103, Asn322, Asn432, Asn546) and one FA2 glycan (blue) is bound to RBD (at Asn343) and the other is that the six ACE2 glycans are replaced by bi-antennary complex N-glycan (FA2)
The result is as follows. When ACE2 glycans are MAN9, the binding energy decreased by 14.7%, and when ACE2 glycans are FA2, the binding energy increased by 9.1%.
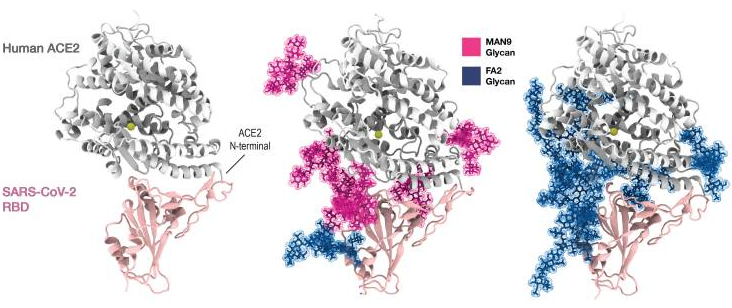
These simulation results seem not to contradict with the other experimental result.
In addition, it was shown that N501Y would introduce additional stabilizing interactions with Y41 and K353 of ACE2, which will increase the binding affinity between RBD and ACE2. The same results from different groups are introduced in the past blog.