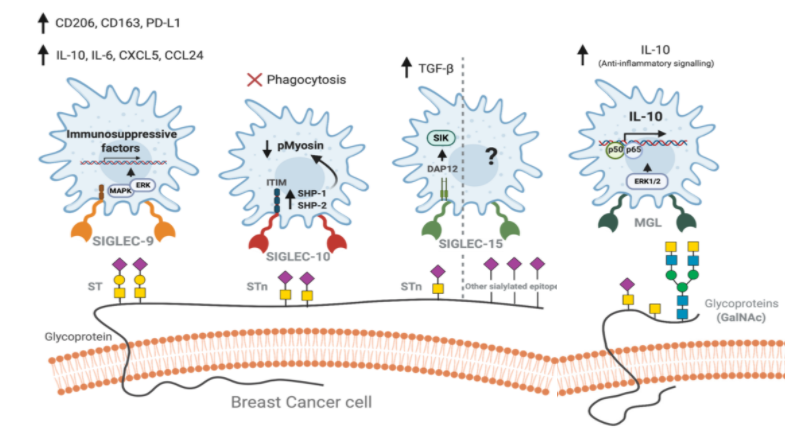
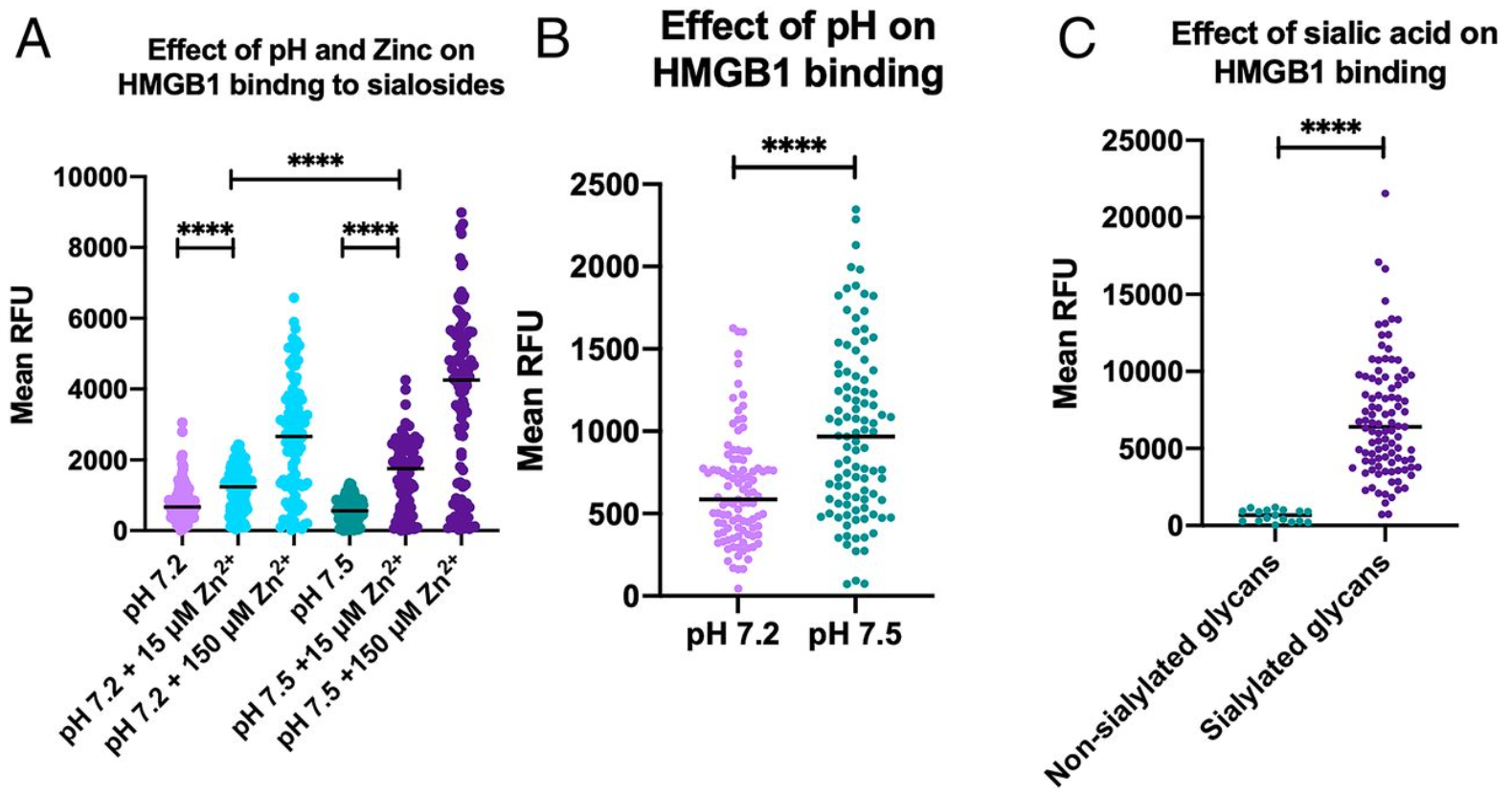
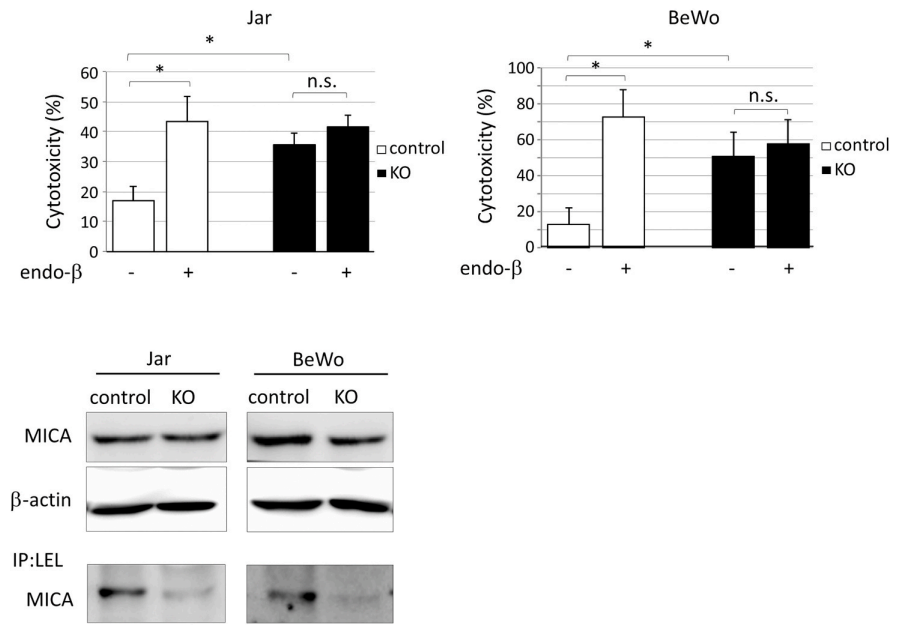
In the new coronavirus (COVID-19), there is a broad spectrum of disease severity from asymptomatic to severe. How the immune reaction is different in the disease spectrum? There are so many studies are ongoing in the world to uncover this problem. A group from Third Military Medical University, Chongqing, China, etc. has reported there is a clear difference in the balance of humoral immunity and cellular immunity between asymptomatic/mild and moderate/severe.
https://www.nature.com/articles/s41392-021-00525-3
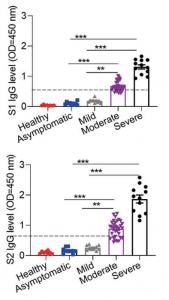
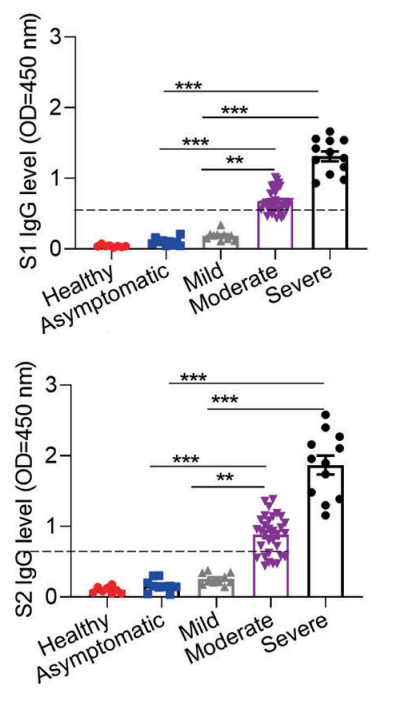
Let’s classify disease severity into two categories, asymptomatic/mild and moderate/severe. In the case of asymptomatic/mild, the expression of SARS-CoV-2 specific IgG is weak and transiently terminated without developing germinal center. In contrast, in the case of moderate/sever, the expression of SARS-CoV-2 specific IgG increases greatly and circulating cTFH cells and chemokine CXCL13, which indicate the magnitude of ongoing germinal center, are upregulated, suggesting that the humoral immunity is strongly activated.
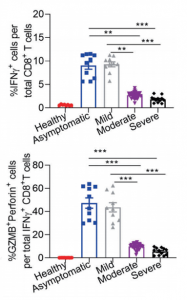
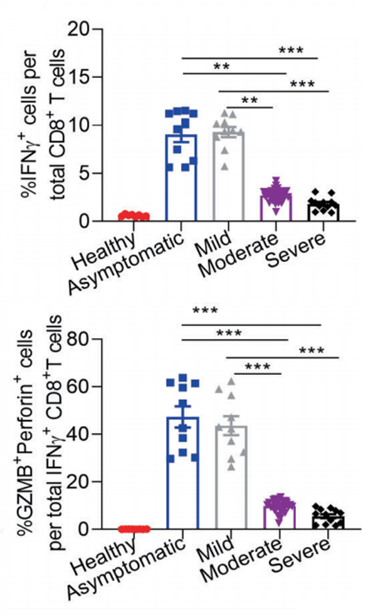
On the other hand, in the case of asymptomatic/mild, IFN-γ producing CD8+ T cells、SARS-CoV-2 specific TH1 cells, Granzyme B(GZMB)are greatly upregulated, suggesting that cellular immunity is strongly activated. (Granzyme B(GZMB)is a Serine Protease expressed on cytoplasmic granules of activated cytotoxic T cells and NK cells, and induce apoptosis of target cells)
That is, asymptomatic/mild is a result of SARS-CoV-2 infection suppression with activate cytotoxic T-cells and NK cells (not mentioned here), and if the viruses are not cleared with this process, the humoral immunity comes to the front to combat viruses, or we can say that it is a result of the balance between humoral immunity and cellular immunity.
Further, recent studies found there exists cross-reactive T cell recognition between circulating “common cold” coronaviruses and SARS-CoV-2I. So, it is of great interest to examine whether the history of “common cold” coronavirus infection with pre-existing SARS-CoV-2 cross-reactive T cells could account for the clinically asymptomatic state in COVID-19 patients.